| Главная |
Введение в природоведческую микробиологию3. ТРАНСПОРТ Транспорт обеспечивает поступление веществ из среды в клетку и выделение продуктов обмена, определяя взаимодействие клетки со средой. В деятельности осмотрофного организма в природе первостепенное значение имеет его способность быстро и до минимальной концентрации использовать субстрат катаболизма из среды. Мембрана бактерий в принципе селективна и может пропускать лишь некоторые вещества, не допуская в то же время выноса веществ из клетки. Разделение зарядов липидной мембраной как основа энергетического обмена и окислительного фосфорилирования сразу же ограничивает возможность проникновения 37 в нее заряженных ионов. Некоторые молекулы поступают в клетку путем пассивной диффузии до выравнивания концентраций. Если в клетке происходит потребление такого вещества, то создается направленный градиент и вещество проходит через липофиль-ную мембрану. Важнейшим веществом, пассивно проходящим через мембрану, является кислород, реагирующий на внутренней стороне мембраны с цитохромоксидазами. Свободно проходят через мембрану молекулы воды, для которых имеются водяные поры, но не растворенные в ней вещества; вода обеспечивает осмотическое состояние клетки. Пассивная диффузия свойственна липофильным соединениям, включая спирты, жирные кислоты. Диффузия важна не только для проникновения веществ в клетку, но и для удаления из клетки продуктов обмена. Скорость диффузии может быть увеличена за счет механизма облегченной диффузии, в котором участвуют пермеазы - пересекающие мембрану белки-переносчики. Связываясь с транспортируемым веществом, они претерпевают конформационные изменения, в результате чего вещество переносится через мембрану в область его более низкой концентрации (рис. 6, 7). Облегченная диффузия ускоряет процесс выравнивания концентраций, но не может привести к концентрированию вещества внутри клетки по сравнению с внешней средой. Трансмембранные белки представляют важнейшее приспособление клетки для взаимодействия с окружающей средой. Они осуществляют не только перенос молекул через мембрану, но и активный транспорт, который позволяет избирательно концентрировать внутри клетки необходимые ей вещества против градиента и, по сути, представляют специфическую химическую реакцию. Активный транспорт осуществляется с затратой энергии 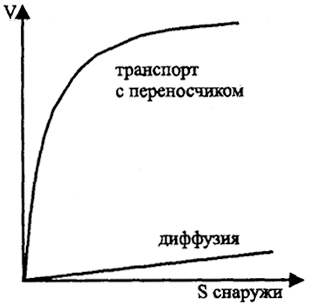 Рис. 6. Поступление веществ в клетку (по Г. Шлегелю, 19.87) S - концентрация субстрата; V - скорость поступления субстрата в клетку 38 либо АТФ, либо за счет протон-движущей силы энергизованной мембраны. От активного транспорта зависит аффинность (сродство) клетки к субстрату - важнейший признак, определяющий и набор, и концентрацию используемых веществ, т.е. зависящую от концентрации кинетику роста. Один из механизмов, транслокация групп, осуществляется при химической модификации переносимого вещества. Лучше всего изучена фосфотрансферазная система Сахаров. Механизм ее действия заключается в фосфорилировании Сахаров на наружной поверхности трансмембранного белка и переносе внутрь фосфорилированного соединения, например, глюкозо-6-фосфата. Фосфотрансферазная система Сахаров представляет собой сложный мультиферментный комплекс, в котором важно отметить два основных компонента: один, ответственный за фосфорилирование данного сахара, а другой (НРr), ответственный за перенос к первому компоненту фосфатной группы, поступающей в конечном итоге от фос-фоенолпирувата. Фосфотрансферазная система используется также при переносе пуринов, некоторых жирных кислот. Перенос через мембрану веществ без их химической модификации, осуществляемый с использованием энергии трансмембранного потенциала, включает три основных механизма: 1) унипорт, когда субстрат (обычно катион, органический или неорганический) транспортируется белком-переносчиком через мембрану, причем другие ионы при этом не переносятся; 2) симпорт, когда транспортный белок обеспечивает перенос через мембрану молекулы субстрата одновременно с переносом в том же направлении протона 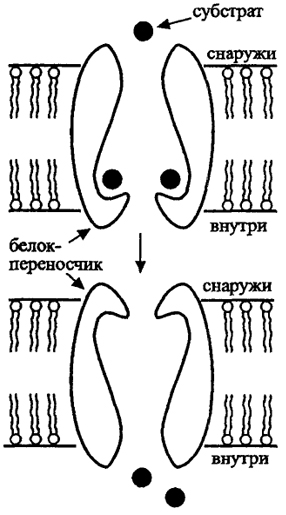 Рис. 7. Перенос вещества через мембрану за счет конформационных изменений белков-переносчиков (по М. Madigan et al., 1997) 39 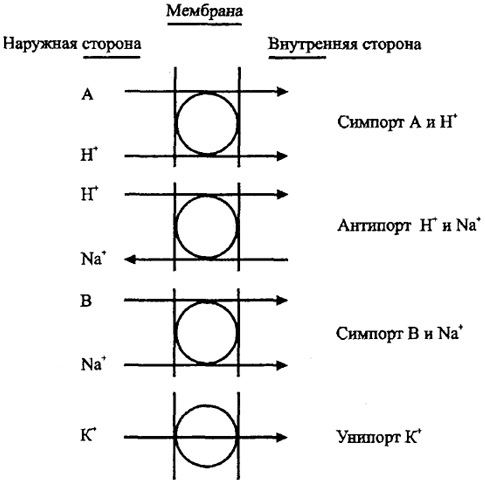 Рис. 8. Активный транспорт за счет энергии протонного потенциала или иного иона; 3) антипорт, когда осуществляется перенос субстрата одновременно с противоположно направленным переносом протона или другого иона (рис. 8). В двух последних случаях молекула транспортируемого субстрата может быть как заряженной, так и незаряженной. Другой пример системы переноса представляет перенос Fe3+ внутрь клетки. В этом случае железо связывается с комплексооб-разующими веществами, сидерофорами, например, гидроксаматом или производными катехина. Образуется комплекс, который и переносится в клетку. Аффинность бактериальной клетки к железу оказывается очень высока. По-видимому, сходные механизмы комплексообразования действуют и для других поливалентных металлов. Специфическая система акцептирования Fe2+ имеется у ацидофильных окисляющих железо литотрофов, как Thioba-cillus ferrooxidans. В этом случае на поверхности клетки есть переносчик - рустицианин, который и вступает в реакцию с электронтранспортной цепью. 40 Перечисленные примеры далеко не исчерпывают разнообразие механизмов транспорта. Следует заметить, что у эукариот разделение клетки на отдельные пространства, ограниченные мембранами (компартментализация), заставляет осуществлять трансмембранный перенос и внутри клетки, что может требовать иных механизмов, чем обмен с внеклеточным пространством. В природоведческой микробиологии особенности транспорта вещества, определяющие набор используемых веществ и сродство клетки к ним, имеют едва ли не большее значение, чем метаболические пути внутри клетки. Особо следует отметить, что клетки обладают разными системами транспорта вещества, включающимися при высокой или низкой концентрации субстрата. Для этих систем транспорта значения Кm (константы, равной половине максимальной скорости реакции) отличаются на 1-3 порядка. 41 |