| Главная |
Введение в природоведческую микробиологию2. КООПЕРАТИВНЫЕ ВЗАИМООТНОШЕНИЯ Кооперативные трофические взаимоотношения в сообществе могут быть рассмотрены на качественном уровне по списку веществ, поступающих в функциональный блок как субстраты, и списку продуктов, образуемых в этом блоке. Наиболее наглядны кооперативные отношения в разнообразной группе органотрофов, осуществляющих деструкционную ветвь в цикле углерода. Осуществляемая микроорганизмами химическая реакция должна обеспечить достаточную энергию для существования организма. Биоэнергетика клетки была разобрана ранее. Здесь же идет речь о взаимодействии видов, осуществляющих разные реакции. В трофической системе микробного сообщества образуется сеть взаимосвязанных химических реакций. Под сообществом мы понимаем набор организмов, занимающих определенные фундаментальные ниши в экосистеме и связанных между собой трофическими взаимоотношениями. Среди этого набора типов взаимоотношений нас интересуют те, которые приводят к формированию микробного сообщества как кооперативного целого, способного взаимодействовать со средой обитания как некое единство. Прежде всего сообщество как целое должно соответствовать требованиям термодинамики и в данных условиях обитания обеспечивать необходимую энергию всем своим компонентам. Для бактерий и их сообществ термодинамика осуществляемых ими химических процессов играет определяющую роль. Далее, из имеющегося набора функционально сходных организмов доминируют те, кинетические характеристики которых более всего соответствуют условиям, складывающимся в сообществе. Сообщество, с химической точки зрения, определяется термодинамикой и кинетикой осуществляемых окислительно-восстановительных реакций. Но физико-химический подход дает только первую приблизительную картину возможностей, 193 реализация которых зависит от биологических особенностей организмов. К таким особенностям относятся, например: способность к выживанию, сопротивление выносу из системы, выеданию и другие свойства, которые могут обеспечить процветание в экологических нишах, входящих в абстрактные "фундаментальные ниши". Микроорганизмы получают энергию для своей жизнедеятельности от окислительно-восстановительных реакций, представляющих перенос электрона от донора к акцептору. По сути дела, в энергетике микроорганизмов мы имеем дело с электрохимической реакцией. Микробиологам такие расчеты особенно удобны для выяснения обмена литотрофных организмов. Общее правило такое: хемолитотрофный организм может развиваться в области термодинамической устойчивости продукта энергодающей реакции. Такие диаграммы приведены, например, в книге Г.А. Заварзина "Литотрофные микроорганизмы" для наиболее известных групп хемолитотрофов (см., например, рис. 3). Однако работает и обратная закономерность: в местообитании сообщества, создающего определенные Eh-pH условия, химические соединения переходят в формы, соответствующие области их устойчивости в так называемых биологически опосредованных реакциях. Для сообщества действует закон Гесса, согласно которому для системы сопряженных реакций важна только разность между исходным и конечным состояниями. При этом допускается множество путей между исходными субстратами и конечными продуктами, лишь бы выход энергии был достаточен. Тот же закон действует и для системы реакций внутри клетки. Однако для сообщества имеется ограничение, заключающееся в том, что для каждого вида, входящего в метаболическую систему сообщества, выход энергии должен быть достаточен для его существования. Таким образом, в многоступенчатой системе реакций деструкции органического вещества каждая ступень, осуществляемая видом микроорганизмов, должна быть выше некоторого минимального уровня. Для того чтобы реакция могла быть энергодающей, во-первых, выход энергии должен быть достаточен для синтеза АТФ, в том числе и за счет протонной помпы, во-вторых, плотность потока вещества должна быть достаточна для поддержания популяции 194 соответствующих организмов, т.е. их урожай У для данной реакции должен обеспечить поддержание популяции организмов вопреки их естественному отмиранию и гибели. Если поток энергии от реакции превращения определенного вещества через локальное местообитание организмов недостаточен для поддержания популяции, то маловероятно и существование специализированной группы организмов. Выход энергии суммарной реакции для сообщества зависит не только от донора электрона, но и от реакций, связанных с окислением. В результате для сообщества деструкторов будут существовать разные возможности, если заключительный этап осуществляется через эндогенные акцепторы, как при брожении или использовании образуемой органотрофами CO2 для метаногенеза или ацетогенеза, или же при использовании внешних доноров, как О2 или SO42-. Поэтому здесь работает обратная связь от конечных этапов к начальным. Пути метаболизма в сообществе, превращающем субстрат s в продукт/? для организмов А, В, С, D... и образуемых ими промежуточных продуктов а, Ь, с, d.., могут быть представлены следующими графами: 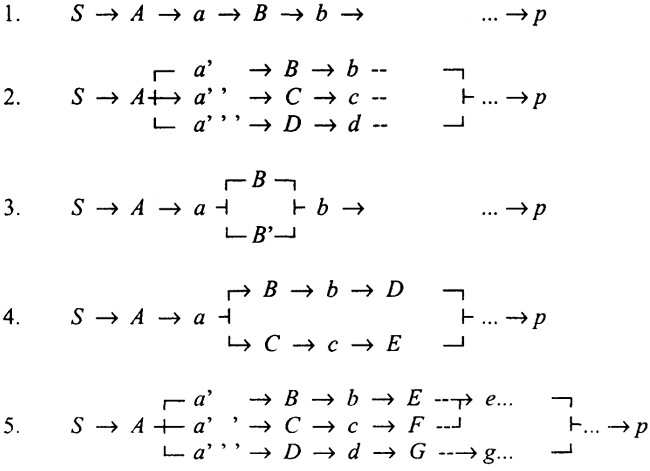 195 На первом графе представлена простая трофическая цепь; на втором - образование разных продуктов, как, например, при смешанном брожении; на третьем - конкуренция за общий субстрат физиологически сходных организмов; на четвертом - конкуренция между организмами, образующими разные продукты; на пятом - комбинация с использованием разных промежуточных продуктов и промежуточным образованием сходных продуктов видами Е и F. Трофическая сеть в сообществе представляет комбинацию разветвленных цепей, ветвящихся либо по продуктам, либо по субстратам. Если р = s, то образуется цикл. Такая структура метаболической сети представляет наиболее общий случай, если рассматривается не только деструкционная ветвь, но и продукционная. В этом случае сообщество имеет внешний источник энергии в виде солнечного излучения, а трофическая цепь относится лишь к материальному балансу. Сообщество, осуществляющее замкнутый цикл (точнее, замкнутые по биогенным элементам циклы) является автономным и способно развиваться в течение длительного, даже в геологических масштабах, времени. Это достигается взаимодействием различных по функциям организмов, составляющих сообщество. При рассмотрении цепи реакций важно заметить, что в условиях сообщества концентрация промежуточных метаболитов а, b... снижается до минимума, обусловленного их потреблением следующим в цепи организмом. Типичным примером может служить низкая концентрация нитрита в реакции нитрификации. 196 |