| Главная |
Эволюционная морфология нервной системы позвоночных§ 1. СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ТАЛАМУСА Таламус, Thalamus, или зрительный бугор, - отдел промежуточного мозга, который претерпевает наибольшие структурные преобразования в ряду позвоночных, что во многом обусловлено его взаимосвязями с прогрессивно развивающимися отделами конечного мозга. Многонаправленность преобразований, недостаточность фактических сведений экспериментальной морфологии, а также неразработанность общих критериев разделения? таламических структур обусловили огромное разнообразие классификаций, а также противоречивость мнений относительно гомологичности отдельных структур таламуса. Строение таламуса низших позвоночных Традиционно в пределах таламуса всех позвоночных выделяют дорсальный и вентральный отделы (ThD, ThV). У круглоротых в состав дорсального таламуса входят расположенные перивентрикулярно слоистое серое вещество, в последнее время обозначаемое как дорсальное таламическое ядро (Thd), а также наружное коленчатое тело (CGL), в состав которого включены описанные ранее как самостоятельные ядро Беллончи; (nВ), переднее и заднее коленчатые ядра. Общими для всех названных образований являются ретинальные входы. Кроме того, пВ получает проекции обонятельных отделов конечного мозга, гипоталамуса и тектума. К переднему коленчатому ядру подходят спинно- и бульбо-таламические тракты, к заднему - проекций октавного комплекса. Эфференты связывают дорсальный таламус с продолговатым и средним мозгом (тегментальная область) и с гипоталамусом. Показан также восходящий путь- tr. thalamofrontalis, - возможно, адресованный гиппокампу. Вентральный таламус у миног отделен от дорсального разреженной зоной (см. рис. 29) и состоит из вентрального таламического ядра, nucl. ventralis thalami (Thv), и ядра заднего бугорка, nucl. tuberculi posterioris (TP). Эфференты последнего, присоединяясь к медиальному продольному пучку, направляются в спинной мозг. Небольшое количество волокон происходит из вентрального ядра. У хрящевых рыб как в дорсальном, так и в вентральном таламусе выделяют медиальную и латеральную части (рис. 65, 66). Медиальная представлена перивентрикулярно расположенными плотно лежащими мелкими нейронами (рис. 65, а), латеральная-мигрировавшими от вентрикулярной стенки, крупными элементами (рис. 65, б). Наиболее крупным отделом дорсального таламуса пластино-жаберных является так называемое латеральное коленчатое тело (ЛКТ). Однако несмотря на идентичность названий доказательства его гомологичности одноименному ядру млекопитающих отсутствуют. В связи с этим ряд авторов предпочитают использовать 200 иное название, другие рассматривают данную структуру как комплекс, состоящий из нескольких отделов. Так, у Ginglymostoma лишь дорсальную часть "ЛКТ" описывают как зрительную, сохраняя за ней это название (другие называют эту часть дорсолатеральным оптическим комплексом); вентральную же часть называют вентролатеральным оптическим ядром. При этом вход сетчатки адресован только последнему, а к дорсальной части поступают лишь тектальные афференты. Эфференты обеих нейронных групп направляются в паллиум. Согласно существующим представлениям вентролатеральное оптическое ядро можно рассматривать в качестве гомолога вентральному ЛКТ, а дорсальную группу - как гомологичную комплексу "дорсальное ЛКТ - LP" млекопитающих. 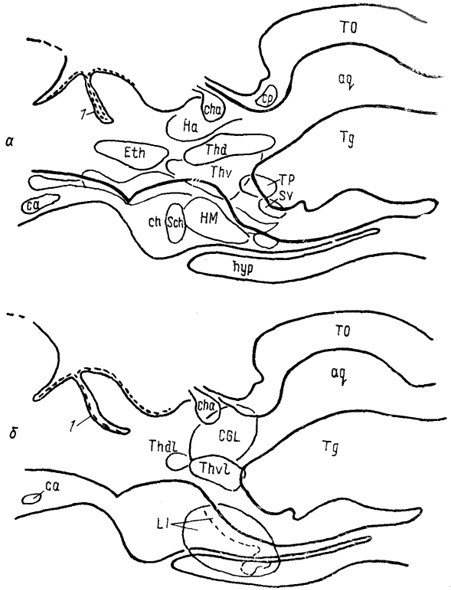 Рис. 65. Отделы промежуточного мозга в проекции на вентрикулярную поверхность III желудочка у акулы Scyliorhinus canicula(Smeets e. a., 1983). а, б - группы ядер: перивентрикулярно лежащие (а), удаленные от вентрикулярной стенки (б). 1 - поперечный парус. Штриховая линия - ядра нижней доли гипоталамуса. Центральное таламическое ядро большинства видов лишено сетчаточных входов, но получает проекции тектума и спинного мозга. Ядро, описанное как латеральное заднее, имеет афференты 201 от мезенцефалического центра представительства электросенсорной системы. Окончания других центров октаво-латеральной системы в таламусе хрящевых рыб не исследованы. Вентральный таламус представлен клеточными массами, формирующими нисходящие проекции к спинному мозгу. Относительно его .афферентов сведения отсутствуют. 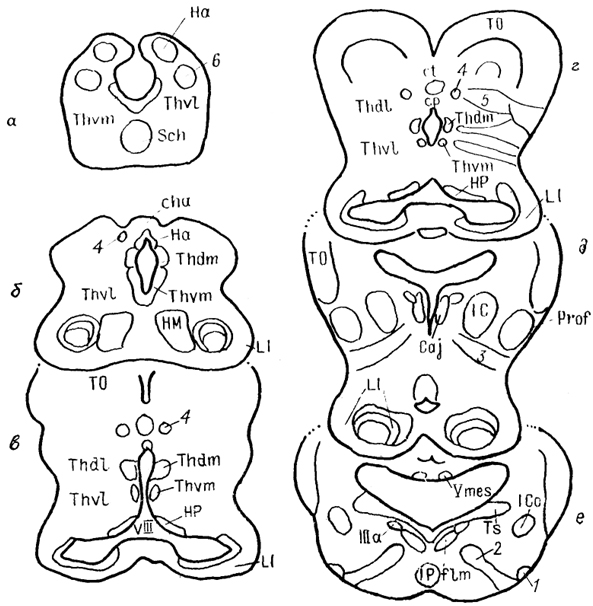 Рис. 66. Строение промежуточного мозга и рострального отдела среднего мозга акулы Scyliorhinus canicula (Reperant e. a., 1986). 1-3 - структуры добавочной оптической системы: базальный зрительный тракт (1), зрительное ядро вентрального тегментума среднего мозга (2), зрительная область, дорсального тегментума среднего мозга (3); 4, 5 - претектальные ядра: оптическое ядро задней комиссуры (4), оптическое центральное ядро (5), 6 - дорсолатеральное ядро, а-e - срезы в ростро-каудальном направлении. В мозге актиноптеригий в составе таламуса помимо дорсального и вентрального отделов в качестве самостоятельной области выделяют задний бугорок, pars tuberculi posterioris (TP) (рис. 67, 1). Он весьма значительно развит у этих животных и вопрос о его гомологичности отделам таламуса других позвоночных не решен. В состав заднего бугорка входит значительное число нейронных групп. В пределах перивентрикулярной области располагаются ядро заднего бугорка, перивентрикулярное 202 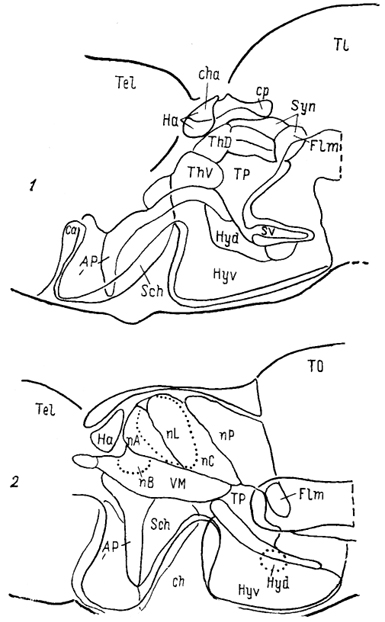
Рис. 67. Схема организации промежуточного мозга представителей многоперовых рыб - Polypterus palmas (1) и бесхвостых амфибий Rana catesbeiana (2) в проекции на вентрикулярную поверхность III желудочка (Braford, Northcutt, 1983). Тонкие линии - контуры ядер, жирные - контуры мозга. ядро заднего бугорка, паравентрикулярный орган и его ядро. Мигрировавшие клеточные массы образуют разное количество групп, среди которых наиболее постоянными являются гломерулярное и прегломерулярное ядра, nucll. glomerulosus et preglomerulosus (Gl, pGl). У большинства представителей ядро Gl - крупная сферическая клеточная масса с четким гломерулярным нейропилем. Ранее оно описано как круглое ядро, nucl. гоlundus, однако используемое в настоящее время название позволяет не только избежать отождествления его с круглым •ядром других позвоночных (что вовсе не подтверждено), но и избавиться от разночтений, поскольку "круглым ядром" разные авторы называли структуры и таламуса и преоптической области лучеперых. Так, ядро G1 отсутствует у карповых, а название "круглое ядро" дано у них структуре, которая значительно отличается от гломерулярного ядра наличием выраженных входов от вторичного вкусового ядра. Прегломерулярное ядро чаще всего рассматривается как комплекс ядер. Дорсальный таламус лучеперых рыб включает перивентрикулярно лежащее переднее ядро, nucl. anterior thalami (nA), a также дорсальное и центральное задние ядра, nucll. dorsalis posterior et centralis posterior (DP, CP). Вентральный таламус состоит из таламического возвышения, eminentia thalami (ETh) и лежащих каудально от него промежуточного, вентролатерального и вентромедиального ядер, соответственно VI, VL, VM (рис. 68). Соотношение основных отделов таламуса у разных 203 групп лучеперых оказывается различным, наибольшие вариации касаются области заднего бугорка. 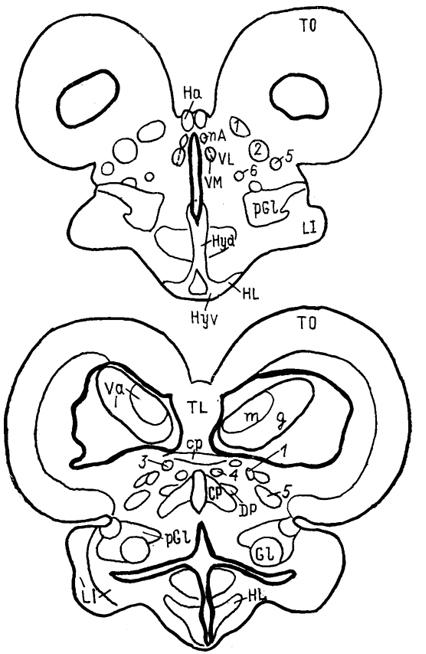
Рис. 68. Ядра промежуточного мозга Carassius auratus (no: Wullimann, Northcutt, 1988). 1-4 - претектальные ядра: центральное (1), поверхностное, крупноклеточная часть (2), 3, 4 - перивентрикулярное: дорсальная (3) и вентральная (4) части; 5, 6 - ядра добавочного зрительного тракта: дорсальное (5) и вентральное (6). Характер связей таламуса лучеперых рыб исследован недостаточно полно. Наиболее многочисленны сведения относительно входов зрительного тракта, которые обнаружены у представителей всех групп. Нейроны, получающие контрлатеральный ретинальный вход, локализованы в пределах переднего таламического ядра, латерально от промежуточного и вблизи вентролатерального. Ипсилатеральные входы организованы различно. Так, у хондростей и голостей они адресованы структурам, имеющим 204 и контрлатеральные входы. На основании сходства в их распределении с таковым у высших позвоночных полагают, что это же было свойственно древним актиноптеригиям, у ранних костистых рыб такие связи были утрачены, а у ныне живущих видов они оказались приобретенными вторично, независимо от других представителей. Действительно, у некоторых телеостей ипсилатеральные входы сетчатки в таламус отсутствуют, у других они возникают в результате двойного перекреста и адресуются ростральным отделам. Сведения о других афферентах весьма фрагментарны. Так, у некоторых исследованных в этом отношении видов, представляющих основные группы лучеперых рыб, обнаружены входы обонятельных луковиц в область заднего бугорка. Тектальные проекции есть у всех ядер, получающих ретинальные афференты, а также у ядер DP, CP, VM; при этом они показаны и у слепых пещерных рыб (в частности, у Astyanax hubbsi - в область "ЛКТ"). Проекции торуса у голостей направляются в ядро СР и прегломерулярный комплекс, однако детали представленности входов от вестибулярного комплекса и органов боковой линии остаются неясными. У костистых рыб проекции боковой линии подходят к области вентрального таламуса, получающей афференты и от латеральной части теленцефалона. Местом окончания церебелло-диенцефальных путей у голостей и телеостей являются ядра VM, nA, TP, при этом основной объем входов адресован VM. Область заднего бугорка получает также восходящие проекции вторичного вкусового ядра и - у телеостей - конечного мозга. Эфференты таламуса лучеперых рыб практически не исследованы. Имеются сведения о восходящих в теленцефалон проекциях от pGl. Дорсальные ядра образуют связи с тектумом, G1- с моторными ядрами V и VII черепномозговых нервов, область заднего бугорка - с долей лицевого нерва. Помимо названных отделов в таламус лучеперых рыб включают так называемый латеральный валик, torus lateralis (Tla). У многоперовых, хондростей и голостей он представляет собой крупное вытянутое скопление нейронов, расположенных на латеральной поверхности в каудальной части промежуточного мозга. У костистых рыб эта структура значительно меньше и смещена рострально. Связи, происхождение и возможные гомологи этой структуры неясны; некоторые авторы считают ее связанной с продольным торусом. Следует отметить, что в числе образований промежуточного мозга у костистых рыб ряд авторов выделяет в качестве самостоятельного еще один отдел - synencephlon (Syn), который занимает пограничную область между промежуточным и средним мозгом (рис. 67, 1). В состав этого отдела включают дорсально расположенную область задней комиссуры, regio com-missurae posterioris, и лежащее вентрально ядро медиального 205 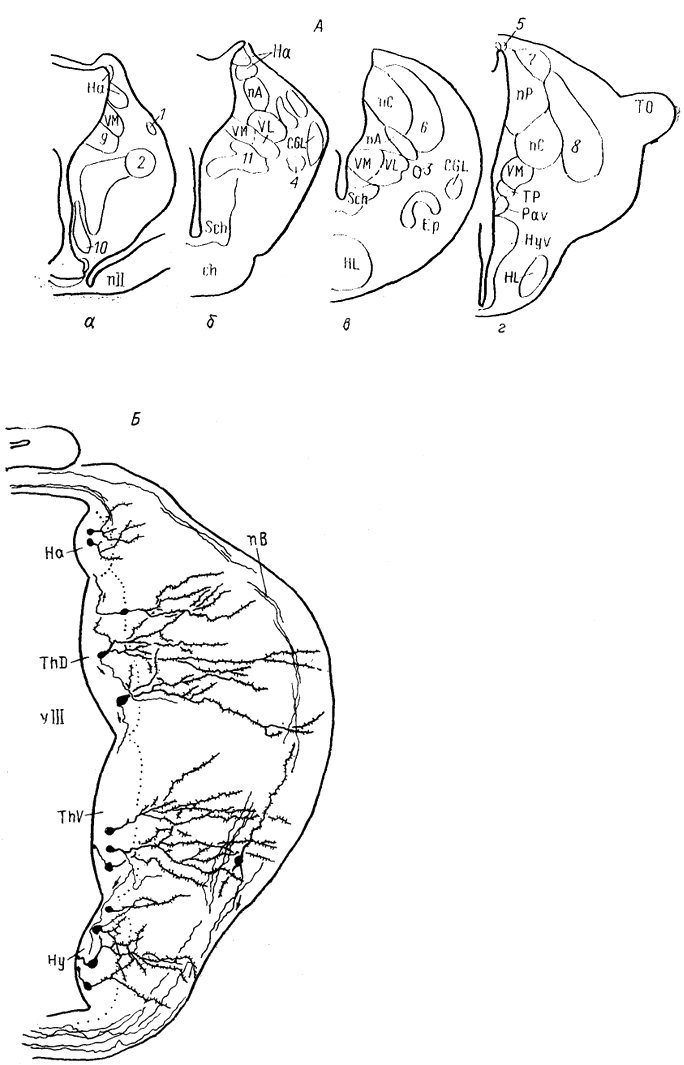 Рис. 69. Схема организации промежуточного мозга представителей бесхвостых амфибий JRana catesbeiana (А) и хвостатых амфибий - Нупоbius (Б) (Neary, Northcutt, 1983; Обухов, 1990). 1 - ядро ложа медуллярной полоски; 2 - латеральная амигдала; 3 - нейропиль Беллончи; 4 - поверхностное желудочковое ядро вентрального таламуса; 5 - субкомиссуральный орган; 6 - 8 - части латерального ядра: передняя (6), дорсальная (7) и вентральная (8); 9-11 - части преоптического ядра; дорсальная (9), вентральная (10) и крупноклеточная (11). а-г - срезы в рострокаудальном направлении. 206 продольного пучка (у большинства позвоночных область задней комиссуры относят к претектальной области). Таким образом, в таламусе водных позвоночных можно выделить, насколько нам позволяют имеющиеся данные, группу ядер, являющихся диенцефальным представительством сенсорных систем. Наибольшее число сведений касается зрительной: системы, однако изучение других сенсорных входов также свидетельствует о наличии на этом уровне центров представительства иных модальностей. Вместе с тем, отсутствие информации о распределении теленцефальных эфферентов и недостаток экспериментально-морфологических исследований не позволяют проводить корректные аналогии с организацией таламуса наземных позвоночных. В мозге амфибий в последнее время также разделяют мигрировавшие и перивентрикулярно расположенные структуры (рис. 69), кроме того, некоторые авторы описывают в пределах таламуса и область заднего бугорка (рис. 67, 2). Дорсальный таламус бесхвостых амфибий состоит из латерального ядра (L) и перивентрикулярно расположенных переднего (nА), центрального (nС) и заднего (nР) ядер (рис. 69, A); двум последним свойственно четкое ламинарное строение. Исходя из характера связей дорсальный таламус делят на три зоны: переднюю, среднюю и заднюю. Передняя зона включает лишь nА. Она получает проекции' от медиального паллиума, вентрального гипоталамуса, энтопедункулярного ядра, области задвижки продолговатого мозга, а" также (за счет ветвления дендритов собственных нейронов в областях окончания ретинальных афферентов и среднемозговом акустическом центре) от структур зрительной и слуховой систем. Входящее в состав передней зоны nА является единственным (по имеющимся на сегодня сведениям) источником таламических входов в медиальный паллиум и вентральную часть дорсального паллиума. Вполне вероятно, что нейроны, дающие начало этим эфферентам, локализованы в одной И той же части ядра, но вопрос о том, являются ли эти клетки источника-ми обоих входов или же те происходят из соседних нейронных групп, остается невыясненным. Отметим, что таламо-теленцефальные проекции, о которых идет речь, билатеральны с преобладанием ипсилатерального звена. Учитывая наличие разно-модальных сенсорных входов в пА, его можно рассматривать как важный (а может быть и единственный) центр проведения сенсорной информации в паллиальные отделы мозга бесхвостых амфибий. Другие эфференты передней зоны адресованы вентральному гипоталамусу, тектуму, задней зоне таламуса. Средняя зона, более крупная, состоит из пС, а также переднего и задневентрального отделов латерального ядра (La, Lpv). Структуры средней зоны получают входы от тектума (La, Lpvy и торуса (nС), кроме того, имеются соматические сенсорные 207 проекции от областей среднего мозга, обладающих соматотопической организацией, и от области задвижки. Основные эфференты этой зоны адресованы стриатуму, и таким образом она рассматривается в качестве важного звена проведения гетеромодальной информации в вентролатеральные области конечного мозга. Небольшое число эфферентов nС направляется в тектум, a Lpv - в торус. Задняя таламическая зона включает в себя ядро nР и заднедорсальную часть латерального ядра, Lpd. Она получает проекции от других диенцефальных областей (передней зоны дорсального таламуса, вентрального гипоталамуса, пинеального органа), стриатума, тектума, кроме того, дендриты ее нейронов, проникая в соседние области, получают ретинальные входы. Эфференты зоны не проецируются в конечный мозг, но адресуются множеству других отделов: моторным ядрам ствола и спинного мозга, тектуму, а также (лишь от ядра пР) торусу, вентральному таламическому ядру VL и nА. Гомологизация ядер дорсального таламуса амфибий с таковыми высших позвоночных не проведена, хотя, как полагают, часть пА, получающая ретинальный вход, может рассматриваться как гомолог дорсального ЛКТ млекопитающих. Вентральный таламус амфибий содержит лежащее перивентрикулярно вентромедиальное ядро (VM) и расположенные латерально ядро Беллончи (nВ), вентролатеральное ядро (VL) и поверхностное желудочковое ядро, nucl. ventricular is superficialis (VS). Кроме того, в составе данной области описано несколько нейропилей, среди которых нейропиль Беллончи и ней-ропиль ЛКТ. Несмотря на наличие значительного ретинального входа пВ не образует проекций в теленцефалон, но направляет эфференты в тектум и заднюю таламическую зону, и на этом основании не может (вопреки ранее существовавшему мнению) рассматриваться как гомолог дорсального ЛКТ высших позвоночных. Дорсальная часть VL в прежних классификациях называлась круглым ядром и относилась к дорсальному таламусу. Однако ни распределение эфферентов, ни характер получаемых афферентов не доказывают гомологичности этой области одноименному ядру рептилий. Для ядер VM и вентральной части VL обнаружены афференты от области задвижки, а для последнего и от сетчатки. Эфференты обоих ядер направляются к тектуму, а ядра VM - и в спинной мозг. Поверхностное желудочковое ядро часто рассматривают как гомологичное вентральному ЛКТ млекопитающих из-за наличия ретинальных входов, однако существующих доказательств этого явно недостаточно. Некоторые авторы полагают, что вся медиальная часть вентрального таламуса амфибий может рассматриваться скорее как гомолог неопределенной зоны промежуточного мозга млекопитающих. В вентральный таламус амфибий часто включают и задний 208 бугорок, представленный небольшой группой перивентрикулярно расположенных нейронов. Однако в отличие от вентрального таламуса эта область формирует проекции в теленцефалон, а в отличие от дорсального - проецируется на спинной мозг. Известные ее афференты происходят из медиального паллиума и стриатума. Такой характер связей не позволяет рассматривать задний бугорок как дериват дорсального или вентрального отделов, но вместе с тем значительно отличает его от одноименной части промежуточного мозга рыб, где основные связи приурочены к ниже расположенным отделам. Строение таламуса высших позвоночных Таламус рептилий весьма сложен и развитие структур его дорсального и вентрального отделов варьирует у представителей разных групп. 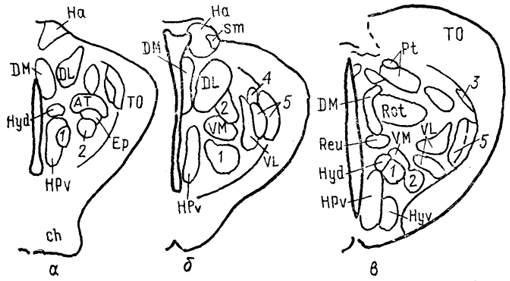 Рис. 70. Строение промежуточного мозга игуаны Iguana iguana (Butler, Northcutt, 1978). 1, 2 - пучки переднего мозга: медиальный (1) и латеральный (2); 3 - лентиформное ядро среднего мозга; 4, 5 - ЛКТ: дорсальное (4) и вентральное (5). а-в - срезы в рострокаудальном направлении. Дорсальный таламус рептилий в ростральной части представлен дорсомедиальным и дорсолатеральным ядрами (DM и DL), в средней части располагаются круглое ядро, nucl. rotundus (Rot), и медиальное ядро, которое у многих смыкается по средней линии и составляет nucl. reuniens (Reu) (рис. 70). Каудальная часть дорсального таламуса включает медиальное заднее ядро (МР). Ядра DM и DL рассматриваются в качестве гомологов ядер передней группы таламуса млекопитающих на основании сходства их эфферентов, направляющихся в стриатум, дорсолатеральный паллиум и гиппокамп. Об их афферентах сведения немногочисленны, имеются указания на тектальные и соматические входы. Ядро Reu получает входы от торуса и представляет собой таламический отдел слуховой сенсорной системы 209 рептилий, его эфференты распределяются в медиальной части дорсального вентрикулярного края и в стриатуме (отметим, что проекции этого ядра исключительно ипсилатеральные). Круглое ядро - одно из наиболее крупных образований дорсального таламуса большинства рептилий (правда, у некоторых: змей оно мало или не описано вовсе). Входы к нему образованы тектумом, дорсолатеральным отделом дорсального вентрикулярного края и стриатумом. Эфференты адресованы латеральной части ДВК. Ядро МР является местом окончания спинно-таламических путей и проецируется на медиальную часть ДВК. (в этом усматривают сходство рептилий с птицами). Вентральный таламус у рептилий включает в себя лежащую за преоптическим углублением area triangularis (AT), энтопедункулярное (Ер), вентролатеральное (VL) и вентромедиальное (VM) ядра, а также овальное ядро (Ov) и латеральное коленчатое тело (CGL). Последнее дифференцируют на дорсальную и вентральную части, каждая из которых получает ретинальные входы. Дорсальная часть обычно рассматривается как происходящая из дорсального таламуса, ее эфференты распределяются в дорсальной коре. Вентральная часть получает также тектальные афференты, а направление ее эфферентов неясно. В составе вентральной части выделяют медиальную клеточную пластинку и латерально расположенный нейропиль. Как тектальные, так и ретинальные афференты билатеральны. Ядра VM и VL часто относят к структурам дорсального таламуса, поскольку (на основании соматосенсорных проекций от спинального лемниска) их гомологизируют с вентробазальньш комплексом млекопитающих. Вместе с тем к этим ядрам, а также к энтопедункулярному подходят проекции от стриатума (от мозжечка - для VL). У млекопитающих такой характер афферентов обнаруживается как в вентральном, так и в дорсальном таламусе, а различия касаются направленности эфферентов. В отношении рептилий сведений для окончательного отнесения ядер к тому или другому отделу недостаточно. Имеются данные о проекциях ядра VL в ростральные области паллиума. Помимо этого у некоторых рептилий оно получает вход сетчатки и проецируется в тектум, что позволило высказать мнение о гомологичности его неопределенной зоне. Ядро VM, как полагают, сходно с субталамическими полями Фореля. Однако обилие вариантов строения затрудняет решение вопросов гомологи-зации даже среди рептилий, а тем более между рептилиями и* птицами, недостаток же фактических данных, в частности сведений об эфферентах, не позволяет провести разделение этих образований в пределах таламуса. В целом в таламусе рептилий возрастает число ядерных образований по сравнению с низшими позвоночными, хотя в структурном отношении сохраняются некоторые примитивные черты организации: существование нейропилей, ветвление дендритов 210 нейронов в пределах других ядер - вероятно-, древний способ обеспечения конвергенции разномодальной информации (рис. 71). Совершенно определенно в таламусе рептилий можно обнаружить выраженную направленность связей в теленцефалон, причем эти связи исключительно ипсилатеральны (в отличие от имеющихся даже у анура). Однако гомологизация структур таламуса рептилий с таламическими ядрами высших оказывается чрезвычайно трудной даже в отношении хорошо изученных отделов зрительной сенсорной системы - число ядер таламуса, получающих ретинальные входы, у рептилий оказалось весьма значительным, особенно у видов с развитым зрением. 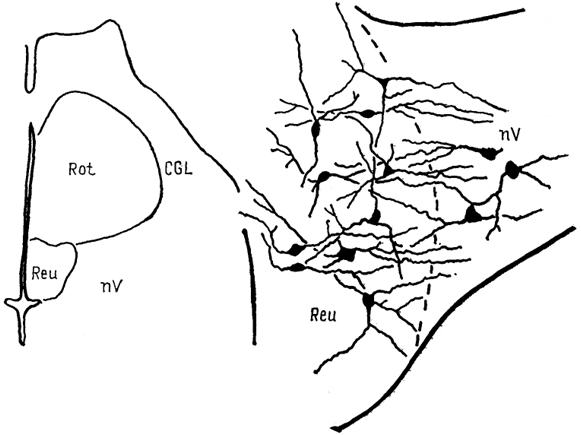 Рис. 71. Нейронные взаимоотношения в таламусе черепахи Emys orbicularis (Белехова, Туманова, 1988). В пределах дорсального таламуса птиц выделяют дорсолатеральное, дорсомедиальное и дорсальное промежуточное ядра (DL, DM, DI), каждое из которых состоит из нескольких групп, обозначаемых как передняя, задняя, медиальная - DLA. DLP, DLM, DMA, DMP и др. и имеющих разную степень развития. Наиболее крупным компонентом таламуса птиц является круглое ядро (Rot). Под ним располагается nucl. triangularis (Tr), вентролатерально лежит прекомиссуральное ядро, nucl. pre-commissuralis principalis, а медиально-задневентральное ядро (VP). Достаточно велико, а у некоторых видов соразмерно с Rot овальное ядро, nucl. ovoidalis (Ov). Его развитие коррелирует со степенью выраженности среднемозгового слухового центра и с развитием слуховой системы. Латеральную часть дорсального таламуса составляет наружное мелкоклеточное ядро, 211 nucl. superficialis parvocellularis (SPC), которое связано с сеп-томаргинальным трактом и получает коллатерали истмо-оптического тракта, а также входы от латерального ядра поводков. Вблизи средней линии на уровне Ov расположена полоска нейронов, обозначаемая как вентральная часть латерального коленчатого ядра (CGLv). Основной частью ЛКТ названо ядро, расположенное вентролатерально от мезенцефалического чечевицеобразного ядра. Несколько нейронных групп лежит в пределах латерального пучка переднего мозга и соединяет диенцефалон с палеостриатумом, с которым они сходны по цитоархитектонике. У некоторых видов часть таких нейронов объединена в энтопедункулярное ядро (Ер). Связи таламических ядер птиц исследованы недостаточно подробно. Известно, однако, что соматосенсорные проекции спинального лемниска адресуются ядрам DLA, DLP, SPC, Rot; последнему адресованы и волокна бульбо-таламического тракта. Волокна зрительного тракта оканчиваются в DLA, а тектальные афференты - в Rot и так называемом nucl. subrotundus (sRot). Ядро Ov, как было упомянуто, получает восходящие проекции среднемозгового слухового центра. Ядро DIP в основном находится под влиянием палеостриатума. Эфференты всех этих образований главным образом направляются в конечный мозг. При этом выделяют ядра дорсального таламуса, которые проецируются исключительно в гиперстриатум - DLA, DLM, DMA, DMP, SPC и DIP. Другие образуют проекции в несколько отделов конечного мозга: в гипер- и неостриатум - DLP, или в экто- и палеостриатум - Тг и Ov. Наконец, sRot устанавливает связи со всеми отделами конечного мозга (рис. 72). Несмотря на имеющиеся сведения относительно распределения эфферентов таламических ядер мозга птиц, далеко не в полной мере удается установить аналоги с функциональными группами ядер млекопитающих. Наиболее четко это можно сделать в отношении сенсорных ядер. Наличие ретинальных входов в DLA и характер его проекций на гиперстриатум, а также ряд других, функциональных, характеристик позволяют рассматривать систему DLA-Wulst как аналог геникуло-стриарной системы млекопитающих. Отметим, что некоторые считают DLA гомологом дорсальной части вентрального ЛКТ. Выраженность тектальных входов, закономерное распределение элементов в соответствии со спектральной чувствительностью и характером ответов приводят к заключению об аналогичности системы Rot-Tr → эктостриатум птиц - экстрагеникулятной зрительной системе млекопитающих, представленной на уровне таламуса комплексом Pul-LP (см. ниже). Однако вопрос о гомо-логичности в данных случаях остается нерешенным в силу недостаточной аргументированности предположений. Остается неясным и положение с аналогами и гомологами системы DLP → промежуточный неостриатум, также имеющей отношение к 212 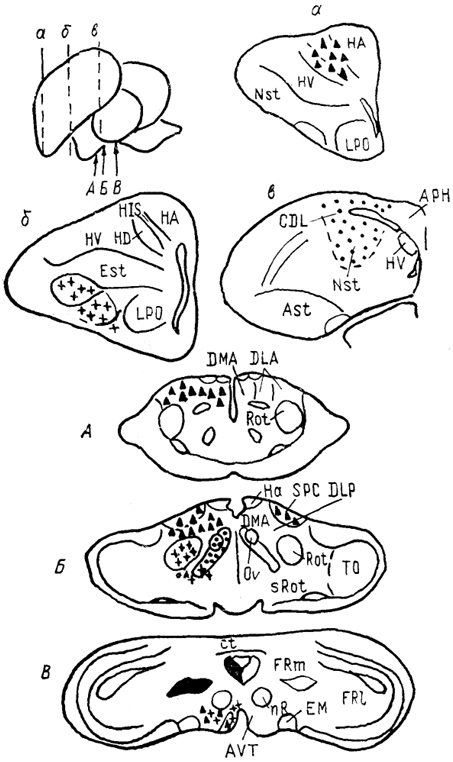 Рис. 72. Схема связей диенцефальных ядер с конечным мозгом птиц (по: Miceli, Reperant, 1985). А-В - уровни сечений промежуточного и а-в - конечного мозга. Одинаковым образом обозначены проекции ядер таламуса в конечный мозг, зачерненные области посылают проекции во все указанные отделы конечного мозга. Обозначение ядер таламуса см. в тексте. осуществлению зрительных функций. Ядра SPC, DLA, DLP, получающие соматосенсорную информацию и проецирующиеся в разные области Wulst, возможно, являются звеньями сомато-сенсорной системы. В то же время систему Rot-Tr часто рассматривают в качестве гомолога вентробазального комплекса 213 млекопитающих. Наконец, ядро Ov, получающее входы нижележащих слуховых центров и по системе ЛППМ проецирующееся в каудальный неостриатум, считают гомологичным ВКТ млекопитающих. Связи ядра Ov с гипоталамусом обеспечивают его участие в процессах вокализации. Однако подобное деление весьма условно и может быть проведено только с целью поиска аналогий в организации систем у птиц и млекопитающих. Так, ядро DLP образует системы проекций к разным областям конечного мозга и, судя по имеющимся данным, с разными функциями (зрительный или соматосенсорный каналы). Ядро DLA, очень напоминающее ЛКТ, получает и волокна спинального лемниска. Оба этих ядра, кроме того, имеют проекции от образований слуховой системы (хотя ответы на звук в них регистрируются с большим латентным периодом). Ядро SPC - со сходными характеристиками, получает также проекции от латерального ядра поводков. Все это может свидетельствовать как о неполноте наших представлений об организации промежуточного мозга птиц, так и о существовании других принципов его строения. Таким образом, попытка применить разработанные для млекопитающих принципы деления ядер таламуса не может быть признана успешной даже в отношении сенсорных таламических образований птиц. Еще сложнее установить аналогии в группах других ядер. Так, Rot получает входы и от претектальной области, и от ретикулярной формации и с учетом этого может рассматриваться как ассоциативное ядро (по имеющимся предположениям, гомологичное комплексу Pul-LP млекопитающих). Если иметь в виду его сенсорные афференты, то аналоги будут иные. Ядро DIP на основании присутствия афферентов от мозжечка и палеостриатума и эфферентов, направленных в Wulst, рассматривается как функциональный аналог ядра VL млекопитающих, получающего проекции базальных гамглиев и лроецирующегося на моторную кору. Остается неясной функциональная роль и, следовательно, аналоги такой системы, как DLM-DMA-"-медиальная часть дорсального гиперстриатума. Особый интерес в сравнительном плане представляет ядро sRot, которое проецируется на гипер-, нео- и палеостриатум. При этом в пределах ядра обнаружены как пространственно разнесенные источники входов к разным областям, так и группы клеток, формирующие за счет коллатералей восходящие проекции к нескольким областям. Такие черты позволяют рассматривать sRot в качестве единственного неспецифического ядра таламуса птиц, хотя среди его афферентов известны лишь входы от тектума. Таламус млекопитающих - наиболее крупный отдел промежуточного мозга, его симметричные отделы соединены у большинства представителей центрально лежащим межбугорным сращением,или massa intermedia. У приматов оно представляет 214 небольшую часть нервной ткани, проходящей через III желудочек, а основная часть таламуса отделена от симметричной половины полостью желудочка на большом протяжении. От окружающих отделов мозга таламус отделен значительным объемом проводящих путей. В каудальной части вентральная поверхность таламуса граничит с субталамусом и гипоталамусом. У большинства млекопитающих каудальная граница наименее четкая - здесь имеется область перекрытия задней группы таламических ядер и претектальной области. Проходящие внутри зрительного бугра внутренние медуллярные полоски (smi) разделяют его на передний, медиальный и латеральный отделы.* Последний содержит вентральную, латеральную и заднюю группы ядер, два других - переднюю и медиальную соответственно. Помимо названных в таламусе млекопитающих выделяют интраламинарные ядра, которые составлены нейронами, лежащими в пределах smi, ядра средней линии, расположенные перивентрикулярно и в пределах межбугорного сращения. Латеральную часть таламуса составляет ретикулярное ядро, nucl. reticularis thalami (Ret), которое обычно не относят к какой-либо из групп. Наконец, у млекопитающих в качестве самостоятельного отдела таламуса часто выделяют так называемый метаталамус, metathalamus, составленный медиальным и латеральным коленчатыми телами (CGL, СОМ). Помимо анатомического деления ядра таламуса млекопитающих часто дифференцируют согласно их функциональной роли II характеру организации проекций: проекционные, ассоциативные и неспецифические ядра. Группу проекционных в свою очередь подразделяют на моторные, сенсорные и лимбические ядра, что отражает их причастность к соответствующим системам мозга и указывает на направленность эфферентов в одноименные области новой коры. Группа ассоциативных ядер характеризуется наличием афферентов от относительно большого числа структур, как стволовых, так и таламических, и распределением проекций в ассоциативных областях коры. Наконец, неспецифические ядра составляют наименее изученную группу, которая традиционно рассматривалась как система со значительным уровнем конвергенции афферентов и диффузным распределением проекций почти по всей поверхности новой коры (последнее, по крайней мере, по современным данным, должно быть подвергнуто пересмотру). Отметим, что такая классификация охватывает лишь таламические ядра, имеющие проекции на области новой коры, и поэтому не может рассматриваться как универсальная. Передняя группа составлена из передних вентрального, медиального 215 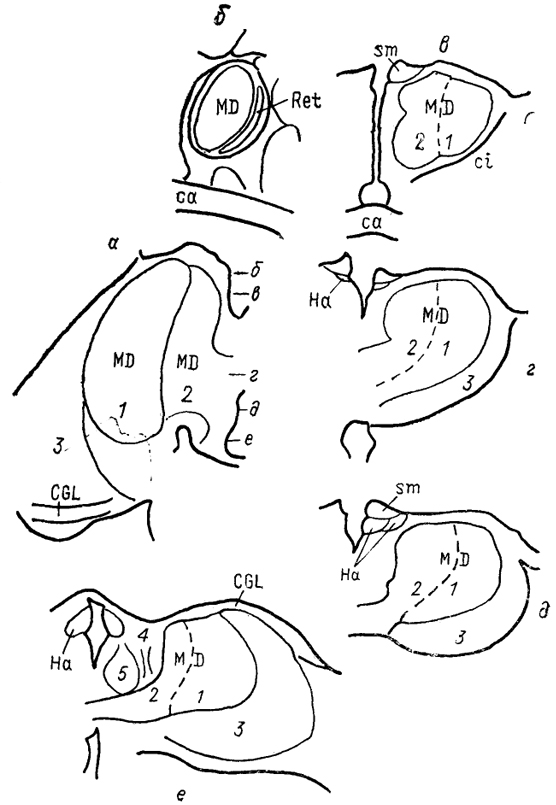 Рис. 73. Таламус ехидны Tachyglossus aculeatus (Regidor, Divac, 1987). a - горизонтальное сечение, б-е - фронтальные сечения на соответствующих уровнях. 1, 2 - части дорсомедиального ядра: крупно- (1) и мелкоклеточная (2); 3 - задневентральный отдел; 4 - хабенуло-интерпедункулярный тракт; 5 - парафасцикулярное ядро. и дорсального ядер (AV, AM и AD) (рис. 74), размеры и относительное положение которых различно у разных представителей. Так, у низших приматов наиболее крупным является ядро AD, у высших приматов и у грызунов, напротив, - это самое мелкое из ядер передней группы, которое зачастую располагается вентрально по отношению к другим. Характерными для ядер данной группы являются афференты от гиппокампа, мамиллярных тел гипоталамуса, цингулярной коры и эфференты, 216 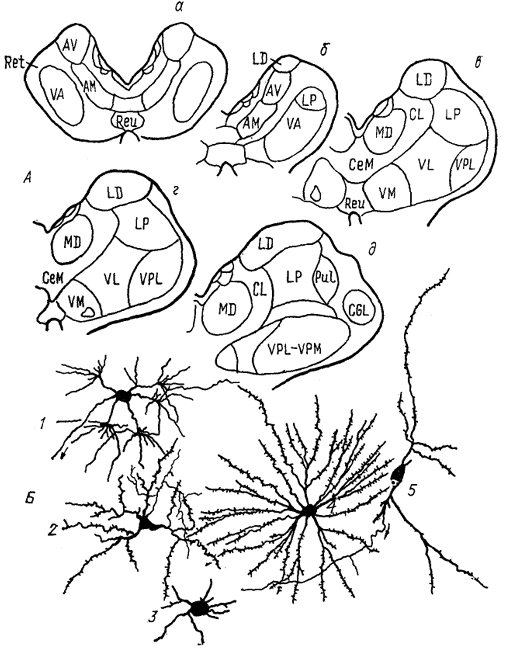 Рис. 74 Таламус кошки (Леонтович, 1979; Macchi, Bentivoglio, 1986). А - расположение ядер на срезах в рострокаудальном направлении (а-д); Б - типы; клеток: 1 - кисточковый, 2, 3 - короткоаксонные, 4 - густоветвистый и 5 - ретикулярный нейроны. Обозначение ядер таламуса см. в тексте. распределяющиеся в области поясной извилины и пресубикулума - на этом основании все ядра передней группы относят к лимбическим проекционным. Не совсем ясен вопрос о наличии этих ядер у некоторых примитивных млекопитающих. Так, у ехидны, согласно современным данным, передний полюс таламуса представлен ядром, относящимся к медиальной группе. Основную часть медиальной группы составляет медиодорсальное (MD) ядро, которое у большинства млекопитающих имеет несколько клеточных групп (рис. 73). Это ядро относится к числу наиболее интенсивно преобразующихся в ходе развития млекопитающих и значительно меняется, например, в ряду приматов - от небольшой гомогенной группы клеток у примитивных обезьян до сложно структурированного крупного ядра 217 у высших приматов и человека. Афференты данного ядра про-.исходят из разных образований мозга, в том числе ядер шва, обонятельного бугорка, амигдалы, гипоталамуса. Эфференты направляются во фронтальную область новой коры, формируя таламо-фронтальную ассоциативную систему мозга. Латеральный отдел таламуса - наиболее крупный у млекопитающих. Вентральная группа ядер этого отдела объединяет значительную часть дорсального таламуса и состоит из многих ядер, часть которых обнаруживается у всех млекопитающих, другие же - только у определенных. К основным в этой группе ядер относят: вентромедиальное и вентролатеральное (VL, VM) -у низших, а у приматов - VL и VA (вентральное переднее), а также заднелатеральное и заднемедиальное вентральные ядра (VPL и VPM). Ядра VM, VL и гомологичные им у приматов VA, VL получают проекции от ретикулярной части черной субстанции, внутреннего сегмента бледного шара, ядер мозжечка и на этом основании, а также с учетом их выходов на неокортекс традиционно причисляются к группе моторных -таламических ядер. Однако лишь ядро VL проецируется в сенсомоторную область коры (MI и SI у крыс, поля 4, 6 у приматов) и таким образом опосредует передачу информации от базальных ганглиев и мозжечка кортикальному звену моторных систем. Что же касается двух других ядер, то их эфференты имеют ряд особенностей: проекции ядра VM широко распределяются по поверхности неокортекса, адресуясь слою I, а ядро VA имеет значительную область перекрытия с ядром MD. Ядра VPL и VPM связаны с проведением соматосенсорной информации, и уровень их развития зависит от степени специализации периферических отделов (развитие лицевой мускулатуры, наличие вибрисс и т. п.). Источниками афферентов этих ядер служат стволовые и спинальные уровни, от которых по многочисленным трактам (спинальный и тригеминальный лемниски, медиальная петля, цервикальный тракт и т. д.) информация поступает в определенные отделы ядер. Таким образом, соматотопический принцип организации нижележащих центров сохраняется и на таламическом уровне: область головы и шеи в основном проецируется в ядро VPM, туловищный отдел, конечности, область хвоста - в VPL. Эфференты ядер направляются в coматосенсорную область новой коры. У некоторых млекопитающих (кошка) в пределах ядра VPM, у других в качестве самостоятельного ядра (вентральное заднее мелкоклеточное - крыса) описана группа клеток, получающая основные входы от парабрахиальных ядер ("мостовая вкусовая область") и от ростральной части ядра солитарного тракта. Эфференты этой группы направляются к инсулярной орбитальной коре (крыса). Аналогом этой области у приматов, вероятно, является медиальная мелкоклеточная часть VPM, также связанная с проведением информации об активации вкусовых рецепторов. 218 Кроме перечисленных к вентральному комплексу относят и другие ядра. Так, у многих млекопитающих описано ядро, называемое nucl. gelatinosus (у крыс) или nucl. submedius (у кошек и приматов). Входы к нему образованы спинальным ядром V нерва и спинальным лемниском и, таким образом, оно может быть отнесено к соматосенсорным ядрам таламуса. Его эфференты у крыс распределяются в небольшой области орбитальной префронтальной коры. Другим ядром является нижнее заднее вентральное (VPI), описанное лишь у приматов (сформировавшееся, вероятно, в результате дальнейшего развития комплекса VPL-VPM). Помимо участия в проведении соматосенсорной информации, возможно, именно за счет связей этого ядра информация от вестибулярных стволовых центров проецируется в верхние отделы мозга медиально от слуховой области коры. Латеральная группа ядер - одна из наиболее изменяющихся в ходе развития млекопитающих, поскольку составляющие ее структуры связаны с филогенетически молодыми областями новой коры. В состав группы входят латеральное заднее (LP) и латеральное дорсальное (LD) ядра. У низших млекопитающих границы между ними нечетки, у высших же ядра достигают .значительных размеров и четко дифференцированы. Кроме того, у высших, как полагают, вследствие развития LP появляется и быстро прогрессирует самое молодое из таламических ядер - подушка, pulvinar (Pul).* Ядра LP и LD рассматриваются в связи с формированием сложных взаимодействий соматической и экстероцептивных систем в ходе развития млекопитающих. Афференты к ним в основном поступают из сенсорных таламических ядер и ядер передней группы. Эфференты у приматов адресуются в основном верхней и нижней теменным областям и поясной извилине. Для низших сведения недостаточно полны; так, у крыс показаны входы лишь в затылочную область коры. Ядро Pul у высших приматов представляет собой крупный ядерный комплекс, составляющий каудальный полюс таламуса. У других приматов, если эта структура присутствует, то она неотделима от LP, например, у тупайи (это справедливо и в отношении млекопитающих, не принадлежащих к приматам). В составе Pul выделяют 4 ядра: переднее, медиальное, латеральное и нижнее (Pula, Pulm, Pul1. Puli); последнее в свою очередь разделяется еще на несколько ядер, различающихся по характеру связей. В целом среди афферентов для Pula доминируют соматосенсорные входы, для Pulm характерны двусторонние связи с миндалиной и височной корой, Pul1 и Puli получают значительное число афферентов от структур зрительной системы, 219 включая разные области зрительной коры, передние холмы (и сетчатку - у макак). При этом проекции лишены рети-нотопической организации за исключением центрального ядра Pul, где обнаружено ретинотопически организованное представительство контрлатерального зрительного поля. Основные эфференты Ptil адресованы заднетеменной и затылочно-височной областям коры. Кроме того, Puli и Puli значительное число эфферентов посылают в экстрастриарную кору, что определяет их участие в организации сложных зрительных функций. Другие отделы комплекса, в особенности Pulm, связаны с височными и фронтальными областями коры, чем, вероятно, обеспечивается участие этого комплекса в организации деятельности слуховой системы и, возможно, системы вокализации. Задняя ядерная группа у низших млекопитающих (сумчатые, грызуны) - относительно большая, но слабо дифференцированная область. Ее состав описывают по-разному, что отражает помимо различий в подходах вариабельность этого отдела таламуса. Так, обычно в ее 'пределах описывают ядра оптического тракта, задней комиссуры, претектальное, а также заднее таламическое (Ро) и надколенчатое, nucl. suprageniculatus (Sg). Первые три часто включают в состав претектальной области. Ядро Sg, хорошо развитое у низших приматов, сходно по нейронному составу и функциональным характеристикам с крупноклеточной частью МКТ. Ядро Ро у высших приматов не описано, у других же его часто включают в состав претектальной области, однако показано, что к нему подходят волокна спинно-таламического тракта и, возможно, оно имеет отношение к проведению информации о болевых воздействиях. Кроме того, у грызунов и сумчатых сюда проецируются некоторые области коры (например, у кошки область SIII). Составляющие метаталамус латеральное и медиальное коленчатые тела (ЛКТ и МКТ) оказываются весьма вариабельными как по степени дифференцированности, так и по уровню развития и размерам, что совершенно очевидно связано с особенностями адаптивной эволюции млекопитающих. ЛКТ - наиболее заметно изменяющийся отдел метаталамуса, обычно подразделяется на дорсальную и вентральную части (ЛКТд и ЛКТв). У млекопитающих, не относящихся к приматам, ЛКТд обычно имеет сравнительно простое строение и представлено недифференцированной группой клеток, лежащих на дорсолатеральной поверхности таламуса. Изменение образа жизни млекопитающих привело на уровне приматов к возрастанию роли зрительной системы и оказалось взаимосвязанным с преобразованиями зрительного сенсорного канала, в том числе и в таламическом звене, где основные изменения затронули ЛКТд, а также привели к некоторому уменьшению ЛКТв, которое у приматов называется прегеникулятным ядром. ЛКТд приматов является основной мишенью ретинальных 220 входов и у большинства имеет ламинарное строение (как правило, 5-6 слоев). Основные слои - парные, каждая пара отличается от другой размерами, связями, характеристиками нейронных реакций. Один из слоев пары получает ипси-, а другой - контрлатеральный вход. Такое разделение входов обнаруживается и у других млекопитающих, однако у приматов, а также у млекопитающих тех групп, в которых зрительная система выполняет роль ведущей, обнаруживаются и иные черты усложнения организации, в частности разделение входов от рецепторов разных типов, что может указывать на общее направление эволюционных преобразований у филогенетически разных ветвей (приматы, нелоцирующие летучие мыши, летучие лемуры, древесные землеройки). ЛКТд - основной источник входов в зрительную область коры. ЛКТв значительно отличается от дорсальной части как по строению, так и по характеристике связей. Ряд авторов рассматривают его в качестве производного вентрального таламуса. В числе афферентов этой структуры помимо образований зрительной системы (передние холмы, зрительная область коры, зрительный тракт) описаны значительные входы от претектальной области. Эфференты (в отличие от дорсальной части) не проецируются в зрительную первичную область коры, а направляются к ЛКТд, передним холмам, претектальной области, добавочной зрительной системе, интраламинарным ядрам таламуса, супрахиазматическому ядру гипоталамуса. Полагают, что из-за различия в распределении эфферентов два отдела ЛКТ у высших млекопитающих различаются и по выполняемым функциям: если ЛКТд имеет большое значение для осуществления предметного зрения, процессов зрительного внимания, распознавания и т. д., то ЛКТв необходимо для обеспечения многих зрительно-моторных реакций, включая зрачковый рефлекс, процессы аккомодации. Следует отметить, что у приматов и нелоцирующих летучих мышей ретинальные входы организованы таким образом, что в ЛКТ (как и в передние холмы) приходит информация от обоих глаз, но с учетом закономерностей перекреста в хиазме - лишь от контрлатерального поля зрения. У других же млекопитающих информация поступает от всех отделов сетчатки, и в представительстве доминирует не зрительное поле, а контрлатеральный глаз. МКТ представляет собой слуховой центр таламуса млекопитающих. В нем подобно ЛКТ обнаруживаются структурно и функционально различные отделы, которые составляют либо мелко- и крупноклеточную части МКТ, либо (по другим классификациям, например, у хищных) - дорсальный, вентральный и медиальный отделы. Основной объем связей вентрального (мелкоклеточного) МКТ происходит из нижележащих слуховых центров, а эфференты распределяются главным образом в первичной слуховой области коры. Крупноклеточная часть, или дорсальный 221 отдел, и рассматриваемое зачастую в комплексе с ним супрагеникулятное ядро нельзя отнести к структурам специфического слухового канала, поскольку в числе афферентов значительный объем занимают проекции несенсорных образований: (например, покрышки среднего мозга), а эфференты распределяются в разных областях височной коры за исключением первичной (от крупноклеточной: части МКТ) и в области островка (супрагеникулятное ядро). 
Рис. 75. Схема организации связей интраламинарных ядер таламуса млекопитающих (Bentivoglio e. а.,1988). 1 - интраламинарные ядра; п.4-п.6 - поля коры. Разная толщина стрелок соответствует разному объему проекций. Интраламинарная группа и ядра средней линии отличаются от всех вышеупомянутых как по строению и особенностям нейронов, так и по характеру связей. Вместе с тем; обе эти группы обычно рассматривают в комплексе, поскольку между ними существует значительное сходство и: в морфологическом и, возможно, в функциональном отношении. По крайней мере до сих: пор ядра этих групп (и только их) относят к так называемым неспецифическим ядрам; таламуса. Состав этих групп несколько различается у представителей разных отрядов. Многие из ядер лучше выражены у низших млекопитающих. На этом основании их часто рассматривают как наиболее древнюю часть таламуса - мнение, которое в свете новых данных, касающихся неспецифических ядер, не может расцениваться как бесспорное. Наиболее заметными являются: интраламинарные ядра: парафасцикулярное, nucl. parafascicularis (PaF) и centrum medianum (CeM). Они как единый комплекс хорошо выражены у хищных, насекомоядных, тупайи, грызунов. Однако в ходе прогрессивного развития млекопитающих ядро СеМ отделяется от PaF и превосходит его по размерам у высших приматов. В структурном отношении СеМ также оказывается весьма сложным и соизмеримо с наиболее развитыми ядрами таламуса у антропоидов и человека. Афференты неспецифических ядер происходят из разных отделов мозга, включая как сенсорные ядра ствола и таламуса, так и структуры моторных и интегративных систем (черная субстанция, 222 претектум, мозжечок, ядра шва, ретикулярная формация и др.), а также определенные области новой коры (рис. 75). Эфференты в основном распределяются в стриатуме и в новой коре. Следует отметить, что в отличие от существовавшего ранее представления о диффузном распределении эфферентов, оно оказалось весьма четко структурированным. Так, для определенных ядер или их отделов могут быть обнаружены области преимущественного окончания эфферентов. Когда от одного ядра проекции направляются к разным областям коры, они не организованы посредством коллатералей, а происходят из разных нейронных популяций. Основные области, получающие входы интраламинарных ядер, - моторные и ассоциативные поля, в то же время первичные сенсорные поля почти лишены интраламинарных входов. Распределение в стриатуме также четко организовано, хотя исследовано менее подробно. Наиболее тесными связями с ядрами конечного мозга обладает ядро PaF. Неспецифические ядра часто рассматривают исходя из структурного сходства и результатов физиологических исследований как ростральный отдел ретикулярной формации. Вместе с тем многие особенности этого отдела таламуса оказываются не до конца понятыми. Во всяком случае в организации связей ядер этой группы описаны черты, не свойственные другим таламическим структурам. К ним относятся тесные связи как с базальными ганглиями, так и с неокортексом, что при учете кортикальных входов в интраламинарные ядра может рассматриваться как путь взаимного влияния коры и стриатума. Отметим, что кортикальные афференты происходят из областей, получающих преимущественные входы от данного ядра группы. Другой особенностью является приуроченность окончания неспецифических афферентов коры (происходящих, например, от ядер шва, голубого пятна, моно-аминергических ядер ствола), минующих переключение в таламусе, к тем же зонам, куда приходит основной объем эфферентов интраламинарных ядер - закономерность, функциональная роль которой пока не определена, хотя очевидно, что подобные связи могут объяснить отнесение интраламинарной группы в состав ретикулярной активирующей системы мозга. Наконец, особенностью данной группы ядер является значительное число билатеральных входов, в том числе и от неокортекса. Последнее обстоятельство с учетом тесных двусторонних связей определенного ядра и определенной области коры привлекает внимание исследователей возможностью рассматривать эту особенность как вероятный путь обеспечения межполушарных взаимодействий. В целом можно заключить, что так называемые неспецифические ядра таламуса млекопитающих представляют собой достаточно сложно организованную и далеко не подробно исследованную группу, которая, судя по имеющимся данным, формировалась вместе с неокортикальными отделами и в ходе эволюции преобразовывалась не менее значительно, чем ядра других отделов (возможно и более интенсивно, чем, например, сенсорные ядра, подверженные в основном адаптивным, а не филогенетическим преобразованиям). Ядро, которое у всех млекопитающих рассматривается самостоятельно и не включается ни в анатомические, ни в функциональные группы, носит название ретикулярного ядра таламуса 223 (Ret). Оно располагается латерально и иногда дорсально от остальных ядер таламуса и составлено тонким слоем нейронов (такая форма характеризует ядро практически у всех плацентарных). Его афферентами являются коллатерали волокон, направляющихся от таламуса в кору и от коры - к стволовым отделам; характер влияния афферентов - возбуждающий. Эфференты адресуются ядрам таламуса. При этом существует четкая пространственная разграниченность - определенные области Ret находятся под влиянием определенной области коры и проецируются на совершенно определенное ядро таламуса, оказывая тормозное влияние. Так, например, каудальная часть Ret у приматов получает возбуждающие входы от зрительной области коры (поле 17) и оказывает тормозное воздействие на ЛКТд. С функциональной точки зрения, Ret часто рассматривается как совокупность тормозных интернейронов, а в морфофункциональном отношении его считают аналогом (или продолжением) звеньев латеральной зоны ретикулярной формации. Субталамус, Subthalamus, располагается вентрально от вентральной группы таламуса. В его состав входят несколько отделов, чаще всего представленных небольшими группами клеток, лежащими среди проводящих путей: неопределенная зона, z. incerta, и ее ядра - перипедункулярное ядро, nucl. peripeduncularis, ядра полей Фореля, nucll. campi Foreli, поля Фореля, содержащие пути, которые соединяют таламус, ядра конечного мозга и ствол мозга, а также два наиболее крупных образования - субталамическое ядро, nucl. subthalamicus, и энтопедункулярное ядро, nucl. entopeduncularis. Последнее занимает самое ростральное положение среди субталамических структур и по своему строению более напоминает стриатарные отделы, а не диенцефальные. Оно часто (особенно у высших млекопитающих) описывается в составе ядер конечного мозга в качестве внутреннего сегмента бледного шара. Для всех названных структур общим является преобладание связей с моторными отделами мозга - в основном со структурами экстрапирамидной системы, поэтому их нередко включают в состав моторных систем мозга. Отметим, что ряд авторов предпочитает рассматривать субталамус в качестве вентрального таламуса млекопитающих. Однако если для некоторых отделов (поля Фореля, неопределенная зона) находят гомологи в мозге других высших позвоночных, то для субталамического ядра не описано даже аналогов ни у птиц, ни у рептилий. Кроме того, если под вентральным таламусом понимать часть зрительного бугра, не проецирующуюся на неокортекс, то ряд структур, описанных ранее (ретикулярное ядро и некоторые отделы ЛКТ), также должны быть включены в него. Тем не менее они по современным классификациям отнесены к собственно таламусу (Ret) и метаталамусу (ЛКТ). Таким образом, применительно к млекопитающим предпочитают использовать термины "таламус" и "субталамус" безпроведения 224 аналогий с делением таламуса других позвоночных. Заключая описание таламуса позвоночных, следует отметить, что в ходе прогрессивной эволюции изменяется степень развития и совершенства разных его отделов: существенно уменьшается область, обозначаемая как синенцефалон, редуцируется область заднего бугорка, а области дорсального (особенно) и вентрального таламуса значительно усложняются, сопровождая преобразования паллиальных отделов мозга. Поиски аналогов и гомологов в пределах таламуса проводились многократно, однако вряд ли эти попытки можно считать успешными. Достаточно четко у всех челюстных позвоночных в пределах таламуса выделяются лишь ядра, имеющие отношение к проведению сенсорной информации в теленцефалон. Но недостаток, в первую очередь морфологических сведений, а также несовершенство критериев дифференциации не позволяют считать их гомологичными, хотя об аналогах с функциональной точки зрения можно говорить достаточно обоснованно. Заметим, что сложность сопоставления сенсорных ядер обусловлена также и значительным проявлением экологических особенностей (как, например, при делении ЛКТ позвоночных на дорсальный и вентральный отделы). Кроме того, некоторые ядра, традиционно рассматриваемые как сенсорные, имеют выраженные гетеросенсорные характеристики (nucl. reuniens у рептилий), что может расцениваться как свидетельство древних черт организации и, таким образом, ставит под сомнение корректность подобных сравнений. Еще одной традиционной точкой зрения является утверждение о постеленной редукции вентрального и прогрессивном развитии дорсального таламуса, которое выражается как в усложнении его, так и в возрастании роли последнего. Однако пересмотр разделения таламических образований, проведенный в последние 10-15 лет, привел к "перемещению" ряда отделов из дорсальной части в вентральную и наоборот. Отсутствие общепризнанных критериев разделения таламуса, по-видимому, не дает возможности рассматривать преобразования в таламусе в этом аспекте. Еще более трудными оказываются поиски аналогий с другими известными ядрами таламуса млекопитающих - ассоциативными и неспецифическими: установить гомологии здесь сложно даже среди млекопитающих, что же касается представителей других классов, то судя по имеющимся данным лишь у рептилий и птиц среди таламических появляются ядра, проецирующиеся на несколько областей паллиума и на этом основании сравнимые с интраламинарными ядрами млекопитающих. У птиц такой структурой является nucl. subrotundus, у рептилий - nucl. perirotundus (у черепах - ядро DM). Следует отметить, что если в качестве определяющей характеристики таламических отделов принять их связи с теленцефальными образованиями, то прогрессивные изменения в этом отношении в ряду позвоночных очевидны: возрастание объема связей, освобождение их от контрлатеральных путей (имеющихся у хрящевых рыб и бесхвостых амфибий), а также прогрессивное разделение и уточнение теленцефальных мишеней. Аналогичные процессы, вероятно, имеют место и в таламических афферентах. Эти преобразования, по-видимому, происходят и в пределах каждого класса. По крайней мере среди млекопитающих многие из этих особенностей можно проследить на группе неспецифических ядер, которые проходят путь преобразований, параллельный изменениям кортикальных отделов. Наконец, отметим, что если для таламуса низших позвоночных обычно существование нейронов, проецирующихся на спинной мозг и моторные ядра ствола, то в таламусе высших такие образования в качестве самостоятельных отсутствуют, а структуры, 225 проецирующиеся на ствол мозга, адресуют свои проекции звеньям надсегментарных или высших моторных систем и, как правило, составляют особые отделы - большинство субталамических образований. Относительно общими остаются для всех позвоночных диенцефальные образования, связанные с организацией зрительно-моторных реакций. Они составляют так называемую претектальную область промежуточного мозга (хотя некоторые ее элементы у низших позвоночных рядом авторов включаются в состав синенцефалона). Претектальная область располагается на границе промежуточного и среднего мозга. Она содержит многочисленные мелкие группы клеток, образующие ядра претектальной области, nucll. pretectales (Pt). В функциональном и структурном отношениях они оказываются сходными с ядрами добавочной зрительной системы, сосредоточенными в области покрышки среднего мозга. Таким образом, характеристику этих образований, целесообразно проводить параллельно. Добавочная зрительная система содержит ядра, получающие прямой сетчаточный вход по добавочному (базальному) оптическому тракту. Она представлена у всех челюстноротых несколькими ядрами. Прежде всего это ядро базального оптического тракта, nucl. basalis tr. optici, и зрительное ядро покрышки, nucl. opticus tegmenti (рис. 66, 68). Гомологизация в отношении других образований проведена далеко не полностью. В мозге рептилий выделяют дорсальную зрительную область и оптическое ядро покрышки среднего мозга, area optica tegmenti mesencephali dorsalis et nucl. opticus tegmenti mesencephali. Последнее было описано под разными названиями: зрительное ядро покрышки, ядро базального оптического тракта, эктомамиллярное ядро. У птиц на основании различий в нейронном составе выделяют три ядра: дорсальное, латеральное и собственно ядро базального оптического тракта. У всех млекопитающих в составе добавочной зрительной системы описывают несколько ядер: медиальное, латеральное и дорсальное терминальные, nucll. terminales medialis, lateralis et dorsalis. При этом у примитивных видов максимально развитыми оказываются медиальное терминальное ядро и подходящий к нему нижний пучок добавочной оптической системы (у ехидны это единственный хорошо выраженный компонент системы). У высших наблюдается увеличение других ядер, а у приматов медиальное терминальное ядро и нижний пучок вообще-трудно выделить. Полагают, что медиальное терминальное ядро гомологично собственно ядру базального оптического тракта, дорсальное и латеральное терминальные ядра - соответственна дорсальному и латеральному ядрам базального оптического тракта птиц. Этот вывод сделан на основании анализа физиологических характеристик нейронов. По этим же критериям не 226 которые из ядер оказались весьма близки к структурам претектума, а в отдельных случаях вообще трудно решить вопрос о принадлежности ядра той или иной системе. В составе претектальной области уже у миног выделяют при-мордиальное претектальное ядро. У хрящевых и костных рыб здесь описывают несколько ядер, в том числе ядро задней комиссуры. 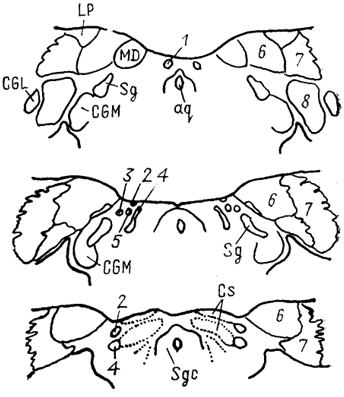
Рис. 76. Претектальная область мозга обезьяны Масаса fascicularis (Kaas, Huerta, 1988). 1-4 - ядра претектальной области: медиальное (1), переднее (2), оливарное (3 и заднее (4); 5 - ядро оптического тракта; 6-8- ядра Pul: медиальное (6), латеральное (7) и нижнее (8). У высших позвоночных состав претектальной области еще более сложен. На основе цитоархитектоники область подразделяется на несколько групп. Так, у рептилий претектальная область весьма значительно варьирует, в ее составе выделяют медиальную и латеральную группы. Латеральная включает в себя три ядра: коленчатое претектальное, nucl. geniculatus pretectalis, чечевицеобразное среднемозговое, nucl. lentiformis mesencephali и заднедорсальное, nucl. posterodorsalis. Медиальная группа называется чечевицеобразным ядром таламуса, nucl. lentiformis thalami, и разделена на перивентрикулярную (медиальную) и латеральную части, pars plicata et pars extensa. Последняя так же, как ядра латеральной группы, значительно преобразуется у некоторых видов с хорошо развитым зрением (агамы, игуаны, тейиды, вараны, хамелеоны) . У этих животных в составе гипертрофированной pars extensa выделяют дорсальное, вентральное и медиальное претектальные ядра. Последнее, присутствующее также в мозге у черепах и кайманов, описано у них под названием дорсального' ядра задней комиссуры, nucl. dorsalis commissurae posterioris. Самым крупным ядром претектума птиц является чечевицеобразное мезенцефалическое ядро, nucl. lentiformis mesencephali, в составе которого выделяются подъядра. Кроме того, к этой области относятся диффузное претектальное ядро, nucl, pretectalis diffusus, претектальная оптическая область, area optiса pretectalis и nucl. synencephalicus superficialis. У большинства млекопитающих в составе претектума выделяют 227 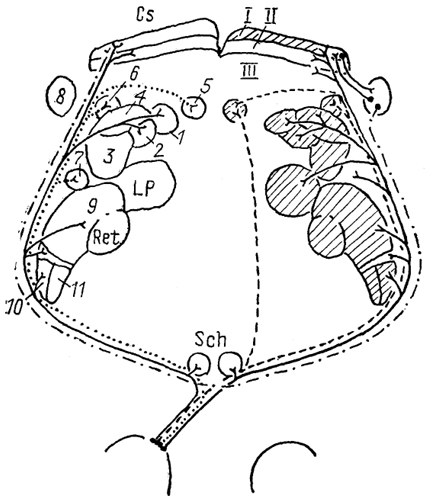 Рис. 77. Схема организации связей структур зрительной системы крысы (Sefton, Dreher, 1985). 1-3 - претектальные ядра: заднее (1), оливарное (2) и переднее (3); 4 - ядро зрительного тракта; 5-7 - терминальные ядра добавочной зрительной системы: медиальное (5), дорсальное (6) и латеральное (7); 8 - nucl. parabigeminalis; 9 - дорсальное ЛКТ. 10-11 - вентральное ЛКТ: крупноклеточная (10) и мелкоклеточная (11) части. I-III - зоны передних холмов: поверхностная (I), зона вхождения зрительного нерва (II), промежуточная (III). Заштрихованы структуры, получающие входы от зрительной области коры. 4-5 ядер. Так, в мозге крыс описывают: ядро оптического тракта, nucl. tracti optici, лежащее среди волокон ручек передних холмов, переднее, заднее и оливарное претектальные ядра, nucll. pretectales anterior, posterior et olivaris (последнее считают гомологом заднедорсального ядра рептилий). Помимо этих ядер в мозге приматов выделяют медиальное претектальное ядро, nucl. pretectalis medialis, нечетко оформленное в мозге грызунов (рис. 76). Полагают, исходя из топографии, хода эмбриогенеза и свойств нейронов, что ядро оптического тракта гомологично чечевицеобразному среднемозговому ядру птиц и рептилий. У ехидны, вероятно, претектальная область представлена группой клеток, лежащей внутри ручек передних холмов и обозначенной как дорсальное терминальное ядро, или дорсальное терминальное ядро оптического тракта. 228 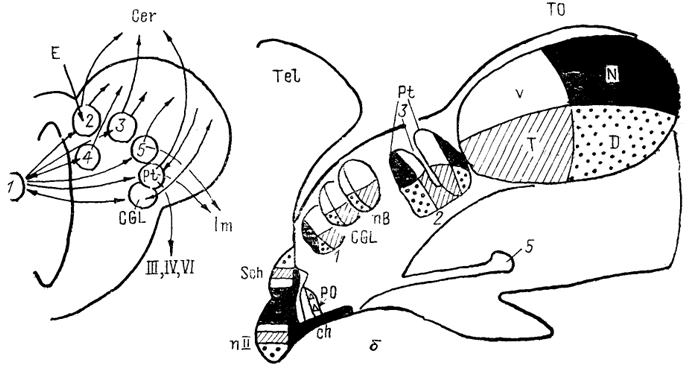 Рис. 78. Схема связей структур, получающих вход сетчатки в мозге костистых рыб (а) и бесхвостых амфибий (б) (по: Vanegas, Ito, 1983; Montgomeri, Fite, 1989). А: 1 - сетчатка, 2 - ядро задней комиссуры, 3 - претектальная область, 4 - дорсолатеральное ядро, 5 - кортикальное ядро, б: 1 - ростральное зрительное ядро, 2 - заднеталамическое ядро, 3 - крючковидный нейропиль, 4 - лентиформное ядро среднего мозга, 5 - ядро базального зрительного тракта. V, N, Т, D - области представительства в контрлатеральной части мозга вентрального (V), назального (N), темпорального (Т) и дорсального (D) квадрантов сетчатки. Характер связей структур претектума и добавочной зрительной системы оказался сходным во многих отношениях. Так, пратически все названные ядра получают более или менее выраженный прямой ретинальный вход (для добавочной зрительной системы преимущественно контрлатеральный, для некоторых ядер претектума -билатеральный). У птиц и млекопитающих во многие ядра обнаружен вход от зрительных областей новой коры (рис. 77) и их аналога в мозге птиц. Наконец, общее заключается в существовании входов от тектума. Сходство их эфферентов заключается в установлении связей с ядрами глазодвигательного комплекса, хотя они выражены по-разному у разных представителей. Так, сложно организованные связи описаны между добавочной зрительной системой и глазодвигательным комплексом у'птиц. Строго упорядоченные проекции имеются также у амфибий, телеостей и рептилий (рис. 78). У млекопитающих не описаны прямые связи добавочной зрительной системы и ядер глазодвигательного комплекса, что же касается претектума, то его структуры проецируются на ядро отводящего нерва (прямо или через медиальную ретикулярную формацию) у низших млекопитающих, у высших же таких связей не обнаружено. 229 Отсутствуют они и у среднемозгового чечевицеобразного ядра птиц. Другое направление эфферентов связывает ядра обеих систем с вестибуло-латеральной долей мозжечка. Причем, у высших позвоночных к прямым входам в мозжечок добавляются и опосредованные нижней оливой, а у млекопитающих описаны исключительно опосредованные (нижней оливой и ядрами моста) связи. У птиц, черепах и некоторых млекопитающих имеются связи добавочной зрительной системы с вестибулярным комплексом, а у птиц и млекопитающих - и с ядрами Кахаля и Даркшевича. Претектальная область таких входов не образует. Анализ распределения связей и изучение характеристик нейронной активности претектальной области и добавочной зрительной системы позволили установить, что эти образования играют решающую роль в организации зрительно-моторных реакций; в частности, опосредуют вертикальный и горизонтальный нистагмы, а кроме того, принимают участие в стабилизационных процессах на сетчатке. Наконец, связи оливарного претектального ядра с ядром Эдингера - Вестфаля обусловливают зрачковый рефлекс. Компоненты обеих систем, имеют тесные связи между собой, а также с тектумом. Однако возможно, что сказанное не в полной мере отражает функциональную роль претектальной области, элементы которой, по крайней мере у птиц, некоторых рептилий и у высших млекопитающих, проецируются на филогенетически молодые таламические структуры и, вероятно, опосредуют проведение зрительной информации в теленцефалон. 230 *
Отметим, что у примитивных млекопитающих таламус часто представлен относительно гомогенной клеточной массой, лишенной внутренних топографических ориентиров, что затрудняет его деление (рис. 73).
*
Отметим, что в составе данной группы у приматов описывают и дополнительные ядра: латеральное промежуточное и, в составе LP, - угловое.
|